

At night they are able to see it in colors, but how?
2013/06/01 Errasti Diaz de Gereñu, Gadea - EHUko biologia lizentziaturako ikaslea | Muñoz Uriarte, Urtzi - EHUko biologia lizentziaturako ikaslea Iturria: Elhuyar aldizkaria

Vision is one of the most important factors in the survival of an individual, essential in the persecution of prey, in the avoidance of predators and in the process of discovery of his partner. It presents a great variability and its ability to adapt to different situations, as well as to its evolution. Thus, some structures of the eye, such as the tapetum luciduma, have become functionally equal in lineages without close evolutionary relationships.
The essential structures to be able to see it are the photoreceptors located in the retina of the eye. They can be cones (color vision) and canes (light intensity). In cones will appear the opsin molecule, which can be of three types: S opsines are sensitive to short-wavelength light, M opsines to intermediate wavelength and L opsines to large wavelengths, allowing to visualize blue-violet, green and red respectively. The types of opsin that each being will have will be genetically determined. In the sticks, instead of opsin, rodopsin appears, which depending on the light intensity of the area will have different level of activity. Despite being inactive in very high illuminations, it is very useful to take advantage of the low existing lighting.
Natural selection, as an evolutionary mechanism, acts differently at every particular moment and place and only the genes that favor over time advance. In origin, the view in the mammal lineage was dicromata (they distinguished two colors), but it has varied depending on the evolution of each evolutionary line, according to the needs of each and the influence of selection. In the original situation, opsina S always appeared and with it opsina M or L. In current cetaceans, for example, the gene encoding opsine S has disappeared and are monochromatic (they see it in a single color range). In the human lineage, on the other hand, in the genes that determined the original dicromata there would have been a duplication and the duplicate gene would have developed a new function, giving rise to the current situation to a trichromata (the three opsines are at once in the retina, being able to distinguish three ranges of colors).
Within visual variability, there are characteristic situations such as the vision model developed in nocturnal animals. For these animals it is very important that the low night light (stars, moon...) is directed to the retina and for this purpose the sticks of the retina are as sensitive as possible. Thus, Solovei et al. discovered in 2009 that, in the clubs of nocturnal animals, the pattern of DNA distribution that appears in the canes of the rest of the animals is inverted, which allows a greater production of rhodopsin, increasing sensitivity to light.

Besides developing the night vision, there are also animals that have developed a more extreme situation: those able to see it at night and of colors. It has always been considered that the visual changes produced during the evolution of mammals are related to the persecution of food and prey. Thus, dicromacy has been related to the animals of the day, while in nocturnal animals it has been considered that color was unimportant. Interestingly, however, current research does not support this idea, as it has been seen that some nocturnal mammals can distinguish colors. Although they do not reach the resolution of the current trichromata, they maintain two opsinas.
In relation to the reasons why the existence of two opsinas has been maintained, the researchers have formulated two hypotheses. According to the first, in these animals dicromacy is not so advantageous in itself, but the plesiomorphological characteristic (inherited from the ancestors) of animals recently converted into nocturnal. In the second hypothesis, dicromacy would be an advantageous feature for nocturnal animals, or the opine genes, in addition to a color-related function, would fulfill some other. Studies show that M and L opsines have remained unchanged in some night lineages compared to their original state, such as aye-aye ( Daubentonia madagascariensis) and some bats.
Aye-ayea
In 2007, Perry et al. They compared the aye-aye DNA sequence with that of other lemuriforms to observe whether or not there was a change in opsin genes and, from there, to respond to the ability to night vision and color. Thus, in the opsinas of aye-ayea no changes were detected with respect to the original dichromatic, and it can be thought that the separation of colors would be an advantage for these nocturnal animals and that the changes produced, if not favored, disappeared.

The aye-ayea, which has long been distributed among the rest of the group, has developed during all this time a series of features to adapt to its ecological niche. However, the opsin genes have maintained their original state over time, reinforcing again the hypothesis about their functionality.
Bats
Bats are divided into two large groups: Fruit megachiropteras ( Yinpterochiroptera) and Microchiropteras ( Yangochiroptera) insectivores have developed different forms of ecolocation, since their needs are different. The former have a good visual and olfactory evolution and the developed eco-location is low-duty-cycle. The sounds they emit are short-lived and do not overlap with echo. In the case of seconds, the echolocation is laryngeal and the waves are emitted constantly, overlapping them with the waves that are returning.
With the different efficiency of both forms of ecolocation, and with less developed eyes on Microchiropter, it is often thought that these bats are based on eco-location and that they do not have very good vision. However, opsin S has remained in both groups, being able to see ultraviolet light and therefore are not blind, as is often said. One of the reasons why bats can see ultraviolet light may be related to the wings of the Gautan lepidoptera that are part of their diet, of which 80% reflect ultraviolet light. On the other hand, another proof of the usefulness of sight is in the hunting form of bats. In fact, after approaching the dam through the waves of echolocation, in the last step they use the view, since being able to detect the waves the insects would be avoided.
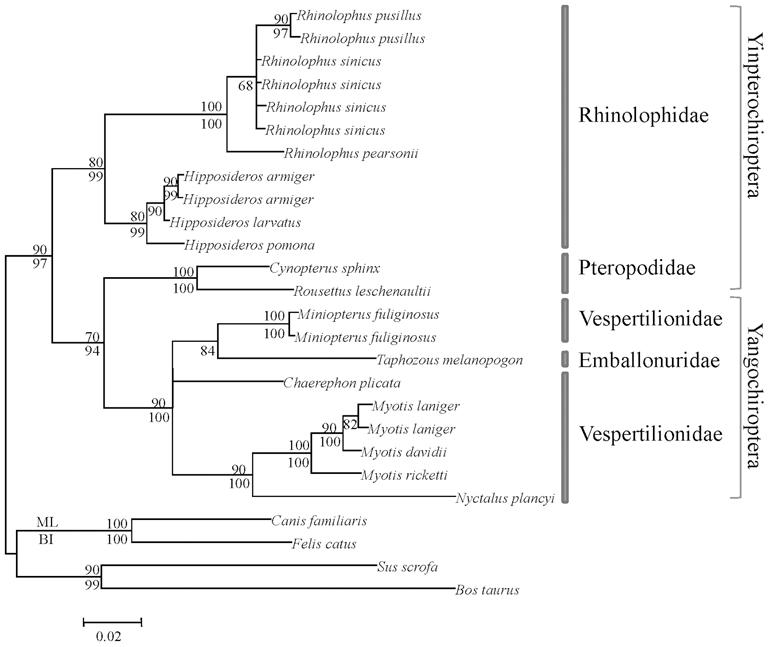
Instead, in Megachiropter, mainly due to its diet, eco-location is less important. To detect when flowers are full of nectars and ripe fruits, smell and sight are fundamental.
According to the results obtained in genetic studies, all groups of bats have maintained opsine S, as well as opsine M or L, highlighting the second. The choice for the maintenance of the L allele may be related to the vision of the red color, indicator of the maturity of many fruits. The phylogenetic trees that are constructed with the data obtained in the studies carried out with these genes also adequately reflect the evolution given in the lineage of bats and confirm the current grouping.
Apparently, the vision has had different forms of development, reaching often convergent situations depending on the needs and etoecological behavior of animals. However, in some nocturnal animals, not mammals and phylogenetically far from them, it has also been found the ability to differentiate the colors at night; the gecko and some sits of the family Sphingidae are able to distinguish 6 colors, such as moonlight or stars. Much remains to be investigated in this field.
References
